Om utviklings-gennettverk støtter den vitenskapelige litteraturen Stephen Meyer
Av Casey Luskin, 25. juli 2024. Oversatt herfra
 I et innlegg i går -lenke så vi at Stephen Meyer skrev mye om Evo-Devo i Darwins Doubt, og effektivt svarer biolog Gerd Müllers foretrukne evolusjonsmodell for hvordan nye kroppsplaner oppstår. Hvis jeg kunne koke ned Meyers argumenter til tre punkter, ville de være:
I et innlegg i går -lenke så vi at Stephen Meyer skrev mye om Evo-Devo i Darwins Doubt, og effektivt svarer biolog Gerd Müllers foretrukne evolusjonsmodell for hvordan nye kroppsplaner oppstår. Hvis jeg kunne koke ned Meyers argumenter til tre punkter, ville de være:
Bilde 1. Trilobit-fossil (Kambrium)
1. Evo-DeVo fokuserer på rollen som spesielle tidligvirkende mutasjoner i utviklingsprosesser har for å generere nye kroppsplaner, men over 100 år med mutagenese-eksperimenter viser at mutasjoner i gener som regulerer utvikling alltid er skadelige (eller i noen tilfeller bare har trivielle effekter). Meyer oppsummerer: "Dette genererer et dilemma: Store endringer er ikke levedyktige; levedyktige endringer er ikke store. I ingen av tilfellene gir den typen mutasjon som faktisk forekommer produserer levedyktige store endringer av den typen som er nødvendige for å bygge nye kroppsplaner. "
2. Vi ser disse skadelige effektene, spesielt i eksperimenter på utviklingsgenregulerings-nettverk (DGRN), komplekse nettverk av gen-interaksjon som regulerer uttrykket av gener tidlig i utviklingen når en organismes kroppsplan begynner å vokse. Etter å ha gjennomgått eksperimentelt arbeid med DGRN, finner Meyer at "Disse DGRN kan ikke variere uten å forårsake katastrofale effekter på organismen."
3. Disse eksperimentelle resultatene på DGRN har dyptgripende implikasjoner for organismens evolusjon, fordi hvis endringer i DGRN er dødelige for et embryo, hvordan kan de da endres til å forklare hvordan nye kroppsplaner utvikler seg? Meyer skriver i boken: "Systemet med genregulering som kontrollerer utvikling av dyre-kroppsplan er utsøkt integrert, slik at betydelige endringer i disse genreguleringsnettverkene uunngåelig skader eller ødelegger det utviklende dyret. Men gitt dette, hvordan kan en ny dyrekroppsplan, og de nye DGRN som er nødvendige for å produsere den, noen gang utvikle seg gradvis via mutasjon og utvalg fra en eksisterende kroppsplan og et sett med DGRN? " (Darwins Doubt, s. 269)
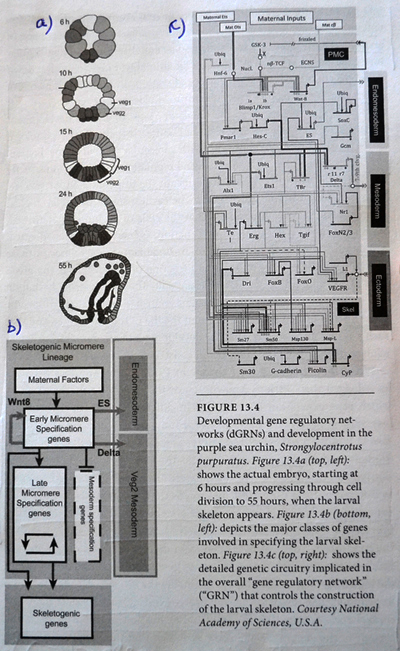 Gerd Müller er klar over at Meyer har snakket om DGRN: - For å komme med det åpenbare argumentet om at mutasjoner i disse genreguleringsnettverkene trenger du ikke så mange tilfeldige mutasjoner for å skape en viktig endring av fenotypen. " Men hvis Meyer har rett, er tilfeldige mutasjoner i DGRN, -dødelige for embryoet.
Gerd Müller er klar over at Meyer har snakket om DGRN: - For å komme med det åpenbare argumentet om at mutasjoner i disse genreguleringsnettverkene trenger du ikke så mange tilfeldige mutasjoner for å skape en viktig endring av fenotypen. " Men hvis Meyer har rett, er tilfeldige mutasjoner i DGRN, -dødelige for embryoet.
Litteraturen støtter Meyers argumenter
Meyer var berettiget til å komme med disse argumentene. Arbeidet med den sene Caltech utviklingsbiolog Eric Davidson, en fremtredende ekspert innen Evo-DeVo, viser at mutasjoner i gener som påvirker kroppsplanegenskaper (som har en tendens til å uttrykkes tidlig, ettersom kroppsplanen blir satt i verk) Ikke fører til nye kroppsplaner - de fører til døde embryoer. Meyer skrev om Davidson i Darwins Doubt, som vi så i går. Men det er verdt å gi litt mer ekspansiv bakgrunn, uttrykt i Davidsons egne ord:
-Det er en høy straff for endring [i DGRN], ved at forstyrrelsen av det dynamiske uttrykket av hvilket som helst av genene, forårsaker kollaps i uttrykket for alle, og det totale tapet fra systemet med deres bidrag til den regulatoriske tilstanden .. Det er alltid en observerbar konsekvens hvis en DGRN -underkrets blir brutt. Siden disse konsekvensene alltid er katastrofalt dårlige, er fleksibiliteten minimal, og siden underkretsene er sammenkoblet, tar hele nettverket i kvaliteten på at det bare er en måte for ting å fungere på. Og embryoene til hver art utvikler seg faktisk på bare en måte.
-For få år siden ble bemerkelsesverdig bevarte underkretser, kalt nettverks "kjerner" som fungerer høyt i DGRN -hierarkiet. ... Kjernene kanaliserer på samme måte nedstrøms utviklingsprosess i hvert medlem for hver gitt gruppe (clade).
[…]
-Evolusjonær mangel på fleksibilitet på grunn av sterkt konserverte kanaliserende DGRN -kjerner
-Som diskutert ovenfor, fungerer disse underkretser på øvre nivåer av DGRN -hierarki for å påvirke karakterene i kroppsplanen som er definitive for taxa på øverste nivå, dvs. de kontrollerer de tidlige stadiene av nettopp de typer utviklingsprosesser, der invariasjonen pr taxa utgjør vårt problem. Siden de utelukker utviklingsalternativer, kan de oppføre seg for å "booleanisere" den evolusjonære selektive prosessen: enten fungerer kroppsdel-spesifikasjonen slik den er ment, eller dyret klarer ikke å generere kroppsdelen og eksisterer ikke.
Eric Davidson, "Evolutionary Bioscience as Regulatory Systems Biology," Developmental Biology, 357: 35-40 (2011
Bilde 2. Oppsett for dGRN (development Gen Regulatory Network)
Eller dette:
-Interferens med uttrykk for [gener i DGRN -kjernen] ved mutasjon eller eksperimentell manipulasjon har alvorlige effekter på utviklingsfasen som de setter i gang. Dette fremhever selektiv bevaring av hele underkretsen, med far for utviklingskatastrofe.
Davidson og Erwin. "Et integrert syn på den prekambiske eumetazoan evolusjonen," Cold Spring Harbor Symposia on Quantitative Biology, 74: 1-16 (2010)
Denne intoleransen for kroppsplan som påvirker DGRN til grunnleggende forstyrrelser indikerer at de ikke kunne ha utviklet seg ved ikke-styrte mutasjoner. Mange koordinerte mutasjoner vil være nødvendig for å konvertere en funksjonell DGRN som genererer en bestemt kroppsplan til en annen DGRN som genererer en annen kroppsplan.
 Det klassiske motargumentet
Det klassiske motargumentet
Meyer er også godt klar over hva Evo-Devo-talsmenn sier som svar på disse argumentene, og han har et klart motsvar. Det klassiske motargumentet fra Evo-Devo-talsmenn er å foreslå at kanskje på en eller annen måte var tidligere DGRN mer 'labile' eller 'fleksible' og lettere for å utvikle seg. Faktisk erkjenner Davidson at noe må ha vært annerledes når kroppsplanene først utviklet seg, noe som fjernet denne motstanden mot endring:
-Å dekonstruere den evolusjonsprosessen som stem-gruppens kroppsplaner trinnvis ble formulert etter, vil kreve at vi krysser den konseptuelle veien til DGRN-eleganse, der ingen moderne DGRN gir en modell. De grunnleggende kontrollfunksjonene til de første DGRN til den prekambiske og tidlige kambriske må ha vært ulike i grunnleggende henseender, fra de som nå blir avslørt i laboratoriene våre. De tidligste var sannsynligvis hierarkisk grunne, snarere enn dype, slik at i begynnelsen kunne adaptiv utvalg fungere i en større del av koblingene. Videre kan vi utlede at utgangene fra underkretsene deres må ha vært polyfunksjonelle i stedet for findelte og funksjonelt dedikerte, som i Modern Crown Group DGRN .. Eric Davidson, "Evolutionary Bioscience as Regulatory Systems Biology," Developmental Biology, februar 2011
Bilde 3. dGRN -tydelige paralleller til kretskort
Davidson sier der at "ingen moderne DGRN gir en modell" for hvordan nye DGRN -er kunne utviklet seg. Derfor mener han at når nye kroppsplaner oppsto, må DGRNs "ha avveket i grunnleggende henseender, fra de som nå blir avdekket i laboratoriene våre." Davidson er ikke den eneste evolusjonære forskeren som bruker denne formen for argumentasjon. Paleontolog Charles Marshall sa omtrent det samme i 2013 da han reagerte i tidsskriftet Science på Meyers argumenter i Darwins Doubt om DGRN. Marshall hevdet at selv om Meyer har rett i å observere at "manipulering av slike nettverk typisk er dødelig," er dette ikke et problem for evolusjonen fordi "GRN -er på tidspunktet for fremveksten av phyla ikke var så kompliserte. Faktisk gir Müllers formidler, Forest Valkai, et lignende (men mindre veltalende uttalt) argument i videoen som angriper Meyer.
 Men hvordan vet Marshall eller Valkai eller noen at DGRN var så forskjellige tidligere? Tilsvarende, hvordan vet Davidson at tidlige DGRN "må ha vært forskjellig i grunnleggende henseender" fra de vi observerer? Vet de dette fra eksperimenter og direkte observasjon, eller fra evolusjonsteorien selv? Svaret er evolusjon; Mer presist: dyrenes felles avstamning. Hvis dyre-phyla delte en felles stamfar som i seg selv var en utviklende art, må DGRN -er fra fortiden ha vært mer "fleksible" eller "labile" - helt ulikt hva vi observerer i dag.
Men hvordan vet Marshall eller Valkai eller noen at DGRN var så forskjellige tidligere? Tilsvarende, hvordan vet Davidson at tidlige DGRN "må ha vært forskjellig i grunnleggende henseender" fra de vi observerer? Vet de dette fra eksperimenter og direkte observasjon, eller fra evolusjonsteorien selv? Svaret er evolusjon; Mer presist: dyrenes felles avstamning. Hvis dyre-phyla delte en felles stamfar som i seg selv var en utviklende art, må DGRN -er fra fortiden ha vært mer "fleksible" eller "labile" - helt ulikt hva vi observerer i dag.
Men ville en så fleksibel eller labil DGRN faktisk produsere et levedyktig dyr? Vi vet ikke, fordi vi ikke har noen observasjonsbevis. Som sådan, for å berge evo-devo-evolusjonsmodeller fra de motsatte eksperimentelle dataene, reverserer Davidson og Marshall den normale metoden for historiske vitenskaper. Nåtidige observasjoner er ikke lenger nøkkelen til fortiden. Snarere dikterer en teoretisk modell hva som må ha skjedd i fortiden - selv om den modellen er i strid med hva vi vet fra bevisene. Meyer uttrykte det på denne måten i epilogen til Darwins Doubt:
Bilde 4. En annen aktuell parallell
-Ved å ignorere dette beviset, reverserer Marshall og andre forsvarere av evolusjonsteori den epistemologiske prioriteringen av den historiske  vitenskapelige metoden som grunnlagt av Charles Lyell, Charles Darwin og andre. I stedet for å behandle vår nåværende eksperimentelt baserte kunnskap som nøkkelen til å evaluere teorienes sannsynlighet om fortiden, bruker Marshall en evolusjonær antagelse om hva som må ha skjedd i fortiden (transmutasjon) for å rettferdiggjøre å se bort fra eksperimentelle observasjoner av hva som forekommer og ikke, i biologiske systemer. Kravene fra evolusjonær lære trumfer dermed våre observasjoner om hvordan natur- og levende organismer faktisk oppfører seg. Det vi vet best fra observasjon, må ty til baksetet, til fordel for tidligere tro på hvordan livet må ha oppstått.
vitenskapelige metoden som grunnlagt av Charles Lyell, Charles Darwin og andre. I stedet for å behandle vår nåværende eksperimentelt baserte kunnskap som nøkkelen til å evaluere teorienes sannsynlighet om fortiden, bruker Marshall en evolusjonær antagelse om hva som må ha skjedd i fortiden (transmutasjon) for å rettferdiggjøre å se bort fra eksperimentelle observasjoner av hva som forekommer og ikke, i biologiske systemer. Kravene fra evolusjonær lære trumfer dermed våre observasjoner om hvordan natur- og levende organismer faktisk oppfører seg. Det vi vet best fra observasjon, må ty til baksetet, til fordel for tidligere tro på hvordan livet må ha oppstått.
-Det vi imidlertid vet av erfaring, er at store økninger i funksjonelt spesifisert informasjon - spesielt informasjon uttrykt i en alfabetisk eller digital form - alltid produseres av bevisste og rasjonelle agenter. Så den beste forklaringen på eksplosjonen av informasjon som er nødvendig for å produsere de kambriske dyrene (enten den eksplosjonen skjedde i løpet av eller før den kambriske perioden) forblir intelligent design. Darwins Doubt, s. 448
Hva dette betyr er at selv om Evo-DeVo har noen interessante ideer, mangler evolusjonsbiologi for øyeblikket en modell som er validert av eksperimentelle bevis som viser at DGRN og derav kroppsplaner-er muterbare og i stand til å utvikle seg fra en form til en annen.
Bilde 5. Med et underliggende naturalistisk prinsipp -er ofte konklusjonen gitt
Flere Evo-Devo-problemer
Men Meyer er ikke ferdig med å fortelle problemer med Evo-devo-baserte evolusjonsmodeller. I Darwins Doubt gir han flere grunner til at mutasjoner i Hox -gener ikke kan bygge nye kroppsstrukturer:
-For det tredje gir Hox -gener bare informasjon for å bygge proteiner som fungerer som brytere som slår andre gener av og på. Genene som de regulerer, inneholder informasjonen for å bygge proteiner som danner delene av andre strukturer og organer. Selve Hox-genene inneholder imidlertid ikke informasjon for å bygge disse strukturelle delene. Med andre ord, mutasjoner i Hox -gener har ikke all genetisk informasjon som er nødvendig for å generere nye vev, organer eller kroppsplaner.
-Likevel argumenterer Schwartz for at biologer kan forklare komplekse strukturer som øyet bare ved å påkalle Hox -mutasjoner alene. Han hevder at "det er homeobox-gener for øyedannelse, og at når en av dem spesielt Rx -genet er aktivert på rett sted og til rett tid har et individ et øye." Han tror også at mutasjoner i Hox -gener hjelper til med å ordne organer for å danne kroppsplaner.
 -I en anmeldelse av Schwartzs bok, finner Eörs Szathmáry Schwartzs resonnement mangelfullt. Han bemerker også at Hox -gener ikke koder for proteinene som kroppsdeler er laget av. Det følger, insisterer han av at mutasjoner i Hoxgener ikke i seg selv bygger nye kroppsdeler eller kroppsplaner. Som han forklarer, "Schwartz ignorerer det faktum at homeobox -gener er valggener. De kan ikke gjøre noe hvis genene som er regulert av dem ikke er der. " Selv om Schwartz sier at han har "undret seg" over "viktigheten av homeobox -gener i å hjelpe oss med å forstå det grunnleggende om evolusjonsendring," tviler Szathmáry at mutasjoner i disse genene har mye kreativ kraft. Etter å ha spurt om Schwartz lykkes med å forklare opprinnelsen til nye livsformer ved å appellere til mutasjoner i Hox -gener, konkluderer Szathmáry, "Jeg er redd for at han generelt ikke gjør det."
-I en anmeldelse av Schwartzs bok, finner Eörs Szathmáry Schwartzs resonnement mangelfullt. Han bemerker også at Hox -gener ikke koder for proteinene som kroppsdeler er laget av. Det følger, insisterer han av at mutasjoner i Hoxgener ikke i seg selv bygger nye kroppsdeler eller kroppsplaner. Som han forklarer, "Schwartz ignorerer det faktum at homeobox -gener er valggener. De kan ikke gjøre noe hvis genene som er regulert av dem ikke er der. " Selv om Schwartz sier at han har "undret seg" over "viktigheten av homeobox -gener i å hjelpe oss med å forstå det grunnleggende om evolusjonsendring," tviler Szathmáry at mutasjoner i disse genene har mye kreativ kraft. Etter å ha spurt om Schwartz lykkes med å forklare opprinnelsen til nye livsformer ved å appellere til mutasjoner i Hox -gener, konkluderer Szathmáry, "Jeg er redd for at han generelt ikke gjør det."
HOX-gener har heller ikke den epigenetiske informasjonen som er nødvendig for dannelse av kroppsplan. Selv i de beste tilfellene endrer mutasjoner i Hox-gener fremdeles bare gener. Mutasjoner i Hox -gener kan bare generere ny genetisk informasjon i DNA. De genererer ikke, og kan ikke, generere epigenetisk informasjon.
Bilde 6. Mutasjoner i hox-gen gir deformasjoner
 I stedet bestemmer epigenetisk informasjon og strukturer faktisk funksjonen til mange Hox -gener, og ikke motsatt vei. Dette kan sees når det samme Hox -genet (som bestemt ved nukleotidsekvenshomologi) regulerer utviklingen av forskjellige anatomiske trekk som finnes i forskjellige phyla. For eksempel, i leddyr, er Hox-genet Distal less nødvendig for normal utvikling av skjøtede leddyrben. Men i virveldyr bygger et homologt gen (f.eks. Dix-genet hos mus) en annen type (ikke-homologt) ben. En annen homolog av det Distal-less genet i echinodermer regulerer utviklingen av rørføtter og ryggraders-anatomiske trekk, som klassisk antas å ikke være homologe med leddyrlemmer, og heller ikke for firefots dyr. I begge tilfeller spiller Distal-less-homologene ulike roller, bestemt av organisme kontekst på høyere nivå. Og siden mutasjoner i Hox-gener ikke endrer epigenetiske kontekster på høyere nivå, kan de ikke forklare opprinnelsen til den nye epigenetiske informasjonen og strukturen som etablerer konteksten og som er nødvendig for å bygge en ny dyrekropps plan. Darwins Doubt, s. 320-321
I stedet bestemmer epigenetisk informasjon og strukturer faktisk funksjonen til mange Hox -gener, og ikke motsatt vei. Dette kan sees når det samme Hox -genet (som bestemt ved nukleotidsekvenshomologi) regulerer utviklingen av forskjellige anatomiske trekk som finnes i forskjellige phyla. For eksempel, i leddyr, er Hox-genet Distal less nødvendig for normal utvikling av skjøtede leddyrben. Men i virveldyr bygger et homologt gen (f.eks. Dix-genet hos mus) en annen type (ikke-homologt) ben. En annen homolog av det Distal-less genet i echinodermer regulerer utviklingen av rørføtter og ryggraders-anatomiske trekk, som klassisk antas å ikke være homologe med leddyrlemmer, og heller ikke for firefots dyr. I begge tilfeller spiller Distal-less-homologene ulike roller, bestemt av organisme kontekst på høyere nivå. Og siden mutasjoner i Hox-gener ikke endrer epigenetiske kontekster på høyere nivå, kan de ikke forklare opprinnelsen til den nye epigenetiske informasjonen og strukturen som etablerer konteksten og som er nødvendig for å bygge en ny dyrekropps plan. Darwins Doubt, s. 320-321
Det vi ser fra passasjen sitert ovenfor, så vel som fra Meyers omfattende diskusjon av Evo-Devo i Darwins Doubt, er at Gerd Müller langt fra har rett i å sammenligne Meyer med en 'genreduksjonist' som mener at bare mutasjoner i gener er nødvendig for å utvikle nye typer organismer. Derimot er Meyer godt klar over ikke-neo-darwinistiske modeller av organisme-evolusjon som Evo-Devo, som fokuserer på mutasjonens rolle i endrede regulatoriske nettverk av gener for radikalt å generere nye kroppsplaner. Meyer undersøker også hvordan ikke-genetisk eller epigenetisk informasjon er nødvendig for å generere nye kroppsplaner, og hvordan evolusjonsmodeller heller ikke virker i stand til å produsere denne informasjonen.
Bilde 7. Epigenenetikk -utgjør en ekstra dimensjon
I sum har Meyer tilbudt omfattende argumenter om Evo-Devo generelt, og spesielt om DGRN. Han viser at DGRN ikke kan endre seg betydelig uten at utviklingen stenger av, noe som betyr at det er et problem med å finne mutasjoner som plutselig kan gi store endringer for å endre kroppsplanen til en organisme radikalt. Siden all evolusjon krever endring, gjelder dette problemet overalt for alle evolusjonære påstander om opprinnelsen til nye kroppsplaner, ikke bare neo-darwinistiske modeller. Joe Rogan-podcasten i fjor ga bare mulighet til å klore på overflaten, men det er tydelig at Meyer har mye å si, og at Müller virkelig ikke har svart ham i noen relevante detaljer.
En invitasjon til dialog
Som et femte og siste poeng vil vi gjerne høre hva professor Müller synes om alt dette. Det er klart at Meyer har investert mye tid og energi i å adressere Müllers felt i Evo-Devo og har utviklet detaljerte, nøye argumenter om levedyktigheten til Evo-Devo-baserte modeller. Jeg ønsker et svar fra professor Müller velkommen. Imidlertid tror jeg at nettstedet vårt, Evolution News, ville være et bedre sted for dialog enn om professor Müllers svarer indirekte gjennom en sint YouTubers kanal, med alle de personlige angrepene og andre usmakelige antikviteter som følger med det aktuelle stedet. Müller fortjener bedre.
Vi vil gjerne være vertskap for en slik dialog her på Evolution News, og jeg inviterer derfor professor Müller til å sende oss et robust svar på Stephen Meyers argumenter om Evo-Devo Modeller om evolusjon. På den måten kan det gjøres virkelig fremgang i denne samtalen.
 Casey Luskin (Bilde 8).
Casey Luskin (Bilde 8).
Casey Luskin er geolog og advokat med høyere grad i vitenskap og jus, som gir ham ekspertise i både de vitenskapelige og juridiske dimensjonene av debatten om evolusjon. Han fikk sin doktorgrad i geologi fra University of Johannesburg, og BS- og MS-grader i geovitenskap fra University of California, San Diego, hvor han studerte evolusjon omfattende, både på hoved- og lavere nivå. Hans jusgrad er fra University of San Diego, hvor han fokuserte studiene på 1.Amendment, utdanningslov og miljørett.
Oversettelse, via google oversetter, og bilder ved Asbjørn E. Lund